A few weeks ago I reported that South Africa had won backing to host the Square Kilometer Array. I got it wrong. The Advisory Committee had recommended that the project go to South Africa, but there was no decision that it would. The final decision is to be made mid-May and apparently the backing of South Africa by the Advisory Committee has not ruled the joint Australia and New Zealand bid out of the race.
Monday, April 16, 2012
Sunday, April 15, 2012
One fish, two fish, red fish, stickleback
Last week an interesting
paper on evolution in sticklebacks, a widespread mostly marine fish, was published in Nature. Sticklebacks are fascinating because populations have repeatedly become established in freshwater at various times since the last ice age. Because of this they provide an amazing system in which the processes of adaptive evolution and speciation can be investigated in the wild.
 |
| The threespine stickleback, Gasterosteus aculeatus. |
Evolution is often defined as the change in allele* frequencies with time. Observations of evolution in the wild, such as previous studies on sticklebacks, show that adaptation to novel environments can happen surprisingly quickly (just 13 generations in one documented case). Too rapidly, some argue, for adaptive alleles to have arisen after the novel environment is encountered. But, the adaptive alleles may already be present in the population at low frequency.
 |
| The left image shows the skeletal differences between marine and freshwater sticklebacks. On the right are preserved specimens stained red. Note the strong divergence in morphology between the marine and freshwater forms, which can arise in just 13 generations (all images David Kingsley). |
If sticklebacks adapted to freshwater through the selection of alleles that arose after they encountered the new environment, then alleles within freshwater populations should be most genetically similar to the populations they diverged form. Conversely, if adaptation occurred through selection on already existing alleles, then freshwater populations should share similar alleles with each other. And the authors tested which of these possibilities was operating in sticklebacks by sequencing the entire genome of ten pairs of sticklebacks. Each pair came from the same area, but one member was the marine form while the other was the freshwater form.
They then examined the genomes for regions that were similar among the marine or among the freshwater fish. Using two different statistical approaches to determine similarities among regions they found 242 regions (0.5% of the total genome) were identified by either test and 147 (0.2% of the total genome) that were identified by both were divergent between marine and freshwater fish. That is, just by looking at those regions in a given fish you could be reasonably confident about whether it lived in marine or freshwater.
Next the authors looked at the genes contained within the regions that were divergent between marine and freshwater sticklebacks. They found that there was a significantly higher density of genes within the identified regions that the genome overall. Then, using the 64 regions that showed the strongest differentiation between marine and freshwater fish, they looked at whether the genes coded for proteins or had a regulatory function. Regulatory genes modify the function of other genes via proteins or RNA.
Regulatory genes were more common within the 64 strongly divergent regions. Just 11 regions (17%) contained coding genes, while 26 regions (41%) were regulatory. The other 27 regions (43%) contained both coding and regulatory sequences, but in these regions none of the changes to coding genes produced different proteins. This strongly suggests that these regions have a principally regulatory effect on trait expression. Thus, regulatory changes account for a much higher proportion of the differences between marine and freshwater sticklebacks.
So, this study is really cool for two reasons. It shows that rapid adaptive evolution to a novel environment can be achieved using the genetic variation present in the parent population. And it shows that the regulation of when and where coding genes are expressed largely accounts for the for the differences between marine and freshwater populations. As the authors acknowledge, the next step is to determine which traits are affected by these genetic differences between marine and freshwater populations.
*Alleles are variants of genes.
Reference (open access!):
Saturday, April 14, 2012
The colour of dinosaurs
When I was young, like many young people, I was obsessed
with dinosaurs. I felt gyped when I found
out that the drawings of dinosaurs in my books weren't based on any real knowledge of dinosaur coloration. The colours used in the dinosaur artwork were just guesses that were extrapolated from the
colours of modern reptiles.
Recently, some of the speculation about the colour of
dinosaurs has been resolved. In the last decade or so there has been an
explosion in the number of dinosaurs that palaeontologists have identified as
having feathers (one spectacular example was revealed just last week). In 2008, a paper1 was published providing details of a
technique that could help to determine the colours of fossil bird feathers. Then,
in early 2010, another paper2 in Nature used the
technique to determine what colour dinosaur feathers might have been.
The structures that give mammalian fur and avian feathers some of their colour are called melanosomes. And, it’s the shape and the arrangement of the melanosomes that help determine the
colour. Excitingly, the 2008 and 2010 papers
showed that these could be preserved in ancient fossil feathers. So, by examining the shape and distribution of the melanosomes in the fossilised
feathers palaeontologists could get and idea of what colour the feathers were.
It didn’t take long for people to start speculating about
the possible function of coloured feathers. Indeed, one of the authors of the 2010 Nature
paper was prepared to speculate when interviewed, that early feathers were for
display and only later were they selected for
insulation and flight. It was, however, only a minor point in their paper. I am skeptical about their claim that display came first, although I agree that selection for flight came much later.
One of the elements that their 'display-first' hypothesis rests on, is that the protofeathers of Sinosauropteryx (the dinosaur they examined melanosomes in) are only present in the tail and in a crest along the dorsal surface of the body. They argue that such limited coverage of proto-feathers suggests that they had a limited role in thermoregulation. But, neither of these points is strictly true.
 |
| A Sinosauropteryx fossil from the Jehol region of China showing the distribution of well preserved protofeathers along the back and tail (figure taken from Chen, P., Dong, Z., and Zhen, S. (1998)3). |
The paper3 that describes the first two specimens of Sinosauropteryx provides evidence that protofeathers were present over most of the body, but were poorly preserved on the animal's sides. Although the protofeathers are most prominent along the back and tail, several small patches were also present elsewhere suggesting a much broader distribution. The paper goes on to argue that the length, density and likely distribution of the protofeathers suggests they would be were most likely for insulation, not display.
Last month a new paper4 provided further evidence of colour in fossil feathers, and speculation about the structure of feathers for flight, display or insulation was given new life. This time there was evidence that the feathers of Microraptor, the stunning four-winged dinosaur, were iridescent. Not strongly iridescent like the throat feathers of hummingbirds or the tail feathers of peacocks, but weakly iridescent like the glossy plumage of crows and ravens.
 |
| An artists reconstruction of Microraptor showing a possible four-winged flight pose for gliding, and it's long midline tail feathers (taken from Li et al. 20124). |
I had been taught that iridescence was all down to the physical structure of the feather, which isn't preserved in fossils, and had nothing to do with melanosome pigmentation, which can be preserved. But, a little reading later and I found that science had moved on! Iridescence requires both the physical structure of the feather and particular arrangements of melanosomes.
As iridescence increases, black melanosomes become arranged in a more orderly way. But, even a well ordered melanosome layer would appear matte black if it wasn't for the structure of a thin keratin layer over the top. Once an ordered layer of melanosomes forms to a thickness of 150 nanometers, no further changes in the melanosomes affect the strength of iridescence, the rest is down to the keratin layer. So, the physical structure of the feather plays the most important role, but an orderly layering of melanosomes is also required5.
The authors of the Microraptor paper were able to infer an iridescent feather colour through two lines of evidence. The shape and the layering of the melanosomes. As far as I am aware, the role that the shape of the melanosome plays in iridescence is unknown. But, in some modern birds the melansomes associated with iridescence have unusual dimensions, and this can be used to distinguish them from other black melanosomes. The authors found that the preserved melanosomes were of the iridescent type and were arranged in a relatively orderly way, strongly suggesting that Microraptor had iridescent plumage.
The authors didn't stop there. The specimen that they had was newly unearthed and provided some interesting additional details about the shape of the tail. It had previously been thought that the tail was quite broad and assisted in flight. But, the new specimen and a re-examination of other specimens, suggested that the tail of Microraptor had two long feathers, or streamers, in the midline right at the back of the tail fan. It also suggested that the tail fan was narrower than previous interpretations.
The authors argue that the iridescent feathers and their new interpretation of the tail shape strongly suggested that the tail was more likely to be for display than to assist with aerodynamics. It's true that in many modern birds iridescent feathers and long tail streamers are important in sexual selection. And it would not be surprising if the tail feathers of Microraptor were used for the same purpose. But, I'm not convinced yet.
When traits are sexually selected, the traits are usually much more exaggerated in one of the sexes. Think the tails of peacocks or the bright colours of male guppies. The other sex is more drab or doesn't have the traits at all. In the Microraptor study, the authors examined three different specimens where the tail feathers were well preserved. All three specimens displayed the elongated tail feathers at the end of the tail.
 |
| Two constructions of Microraptor showing different possible gliding positions. Note the size and arrangement of the tail feathers in comparison to the one above (right image is taken from Xu et al 20036, left image is taken from Chaterjee & Templin 20077). |
When traits are sexually selected, the traits are usually much more exaggerated in one of the sexes. Think the tails of peacocks or the bright colours of male guppies. The other sex is more drab or doesn't have the traits at all. In the Microraptor study, the authors examined three different specimens where the tail feathers were well preserved. All three specimens displayed the elongated tail feathers at the end of the tail.
 |
| Closeups of three fossilised Microraptor tails. A and B are previously described fossils, while C is from the newly described fossil. The arrows in A and B point to the elongated midline tail feathers (taken from the supporting online material of Li et al. 20124). |
Without a clear demonstration of sexual dimorphism for tail
feather length, it's harder to buy the argument that the shape of Microraptor's
tail is sexually selected. And unfeathered tails have recently been shown to be
important for aerodynamics, so I'm not confident that narrowness of the tail
fan rules out an aerodynamic function. The new paper certainly does the sexual
selection argument no harm though. And it shows us another amazing fossil from
China.
References
1 Vinther, J., Brigs, D. E. G., Prum, R. O., and Saranathan,
V. (2008) The colour of fossil feathers. Biology Letters 4, 522 - 525.
2 Zhang, F., Kearns, S. L., Orr, P. J., Benton, M. J., Zhou,
Z., Johnson, D., Xu, X., and Wang X. (2010) Fossilized melanosomes and the
colour of Cretaceous dinosaurs and birds. Nature 463, 1075 - 1078.
3 Chen, P., Dong, Z., and Zhen, S. (1998) An exceptionally
well-preserved theropod dinosaur from the Yixian Formation of China. Nature
391, 147 - 152.
4 Li, Q., Gao, K., Meng, Q., Clarke, J., Shawkey, M.,
D'Alba, L., Pei, R., Ellison, M., Norell, M., & Vinther, J. (2012).
Reconstruction of Microraptor and the Evolution of Iridescent Plumage Science,
335, 1215-1219
5 Maia, R., D'Alba, L., and Shawkey, M. D. (2011) What makes
a feather shine? A nanostructural basis for glossy black colours in feathers.
Proceedings of the Royal Society B 278, 1973 - 1980.
6 Xu, X., Zhou, Z., Wang, X., Kuang, X., Zhang,. F. and Du,
X. (2003) Four-winged dinosaurs from China. Nature 421, 335 - 340.
7 Chatterjee, S. and Templin, R. J. (2007) Biplane wing
planform and flight performance of the feathered dinosaur Microraptor gui.
Proceeding of the National Academy of Sciences 104(5), 1576 - 1580.
Wednesday, April 11, 2012
Older fourlegs
China seems to have had the lion's share of the cool fossils unearthed in the last 10 years or so. A new paper1 on a coelacanth fossil from south China is another example of their phenomenal treasure trove of fossils. It pushes back the origin of anatomically modern coelacanths by 17 million years to 409 million years ago. The previous oldest coelacanth was known from a jaw found in Australia.
Coelacanths are interesting for many reasons, not least because they are more closely related to us than they are to other fish. The group was though to have gone extinct around the same time as the dinosaurs, 65 million years ago. But, in 1938 an extant representative was found in South Africa (although it had been known by the local fishermen for a while before that). A book about the discovery dubbed the fish 'old fourlegs'. There was more excitement in 1997, when a second extant species of coelacanth was found off Indonesia.
 |
| A preserved specimen of the extant coelacanth, Latimeria chalumnae, or old fourlegs. |
The modern coelacanths look almost the same as their ancestors did a few hundred million years ago. So, much like the sharks, they represent 'living fossils'. To put their ancient history in a little perspective, about the same time as the anatomically modern coelacanth body plan emerged, our ancestors were probably taking their first steps on land2.
References:
1 Zhu, M., Yu, X., Lu, J., Qiao, T., Zhao, W., and Jia, L. (2012) Earliest known coelacanth skull extends the range of anatomically modern coelacanths to the Early Devonian. Nature Communications 3, doi: 10.1038/ncomms1764
2 Niedz´wiedzki, G., Szrek, P., Narkiewicz, K., Narkiewicz, M., and Ahlberg, P. E. (2010) Tetrapod trackways from the early Middle Devonian period of Poland. Nature 463, 43 - 48.
References:
1 Zhu, M., Yu, X., Lu, J., Qiao, T., Zhao, W., and Jia, L. (2012) Earliest known coelacanth skull extends the range of anatomically modern coelacanths to the Early Devonian. Nature Communications 3, doi: 10.1038/ncomms1764
2 Niedz´wiedzki, G., Szrek, P., Narkiewicz, K., Narkiewicz, M., and Ahlberg, P. E. (2010) Tetrapod trackways from the early Middle Devonian period of Poland. Nature 463, 43 - 48.
Tuesday, April 10, 2012
XKCD and the ocean
The XKCD webcomic has some nice ocean related comics from time to time. The most recent of which seems to have been inspired by James Cameron's excursion to the Challenger Deep. It gives you a nice feel for just how deep his little submarine took him.
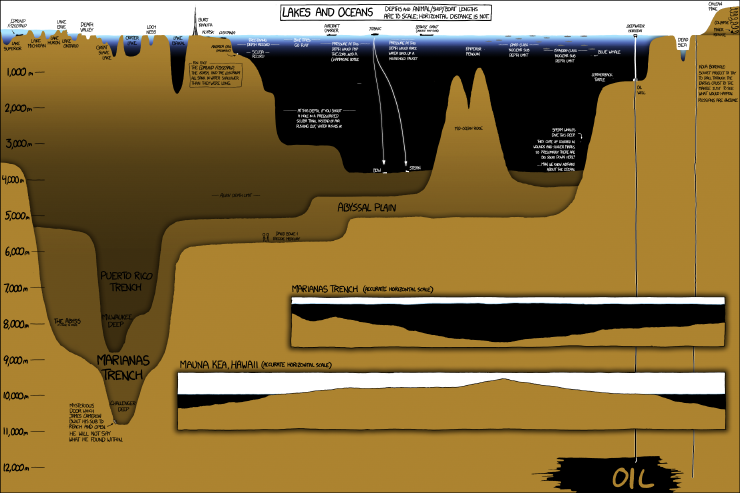
My favourite though, is the one below because it accords with my sense that the surface of the sea is so familiar to us, but we have so little understanding of what's going on beneath.

Monday, April 2, 2012
Bisexual dolphins or media fail?
Total. Media. Fail... Again...
While there is some evidence that dolphins display homosexual and bisexual behaviours, this, this, this, this and many other places are examples of the media completely failing to communicate the results of a scientific study1. The results of the paper in question do not provide any evidence, at all, that would suggest that male dolphins are bisexual. The media has jumped on the word bisexual, which is used just once in the introduction (i.e. before the results of the study are described), and failed to understand it in the context that it was used; perhaps willfully in order to create a more sensational story.
The sentence in the paper reads:
The social system of Indo-Pacific bottlenose dolphins (Tursiops sp.) in Shark Bay, Western Australia, features a fission–fusion grouping pattern with stronger associations between adult males than adult females and bisexual philopatry.
The word 'philopatry' provides the context in which the word 'bisexual' should be interpreted. Here it does not refer to the sexual behaviour of the dolphins, but the fact that both sexes are philopatric. That is, both sexes have the tendency to stay in the area that they were born.
Although I can't be sure, I think the churnalism all started with a story on Discovery News. The story does do a pretty good job of discussing the results of the study, but includes the sentence:
The first order male alliances can also cooperate with other first order groupd to produce a second order alliance, which in turn may join to form a third order alliance. The higher level alliances cooperate to 'steal' females from smaller groups. But, lower level alliances can also recruit other first order groups to defend the female they are herding. If a female is successfully stolen, only one of the joined first level alliances will herd her afterwards.
The higher level alliances are much less stable than first order alliances. For example, groups A and B may cooperate to steal a female from group C one day, then groups B and C may cooperate to steal a female from group A on another day. However, some high level alliances may persist for well over a decade. The reasons for such fluid, reversible alliances at higher levels are unclear, but it may have to do with the context of the encounters.
Despite the findings being less 'juicy' than the popular science articles make it seem, the paper is really interesting. The purpose of the paper was to test hypotheses about the structure of dolphin societies. Evidence from previous studies, such as the one described above, is strongly suggestive that males form their strong alliances in otherwise open social networks. This type of social structure has never been previously documented in mammals.
In mammals, male alliances form to acquire or defend valuable resources from other males, such as territories or female groups. Although it did not seem like this was occurring in the Shark Bay dolphins, two hypotheses had not been properly tested. The 'community defence model', which argues that there are dominant alliances that range over the entire population, and the 'mating season defence model', which argues that males defend territories or females only during the breeding season.
The study found little evidence to support either of the two hypotheses. No male alliances ranged over the entire site as the 'community defence model' predicted. Alliances also overlapped during the mating season and females were not consistently associated with particular alliances as the 'mating season defence model' predicted. So, now we have strong evidence that dolphin societies are unlike any other mammal societies.
The authors of the study point out that a society organised in such a way would put unusual pressures on social cognition. Not only must they remember previous encounters with other dolphins, but they must cope with uncertainty that the relationships are still the same. Individuals that were friends in earlier encounters may have formed new alliances since you last interacted with them and become foes.
It's little wonder that social complexity correlates quite well with brain size, but it's hard to tell which came first. Larger brains may allow more complex social interactions to occur, but more complex social interactions may place selection pressure on large brain size. The authors argue that the complexity of dolphin society is probably a driver of brain size evolution in dolphins, and I tend to agree.
The authors also make the interesting argument that increased locomotion efficiency and low movements speeds have played a role in producing the social complexity and, therefore, brain size evolution. They argue that more efficient travel allows individuals to exploit larger areas and this would increase the chance that competitive encounters occur. A greater number of competitive encounters would, in turn, favour the formation of alliances with other individuals. I'm not so sure about this argument. Brains are energetically expensive to maintain, and probably favour more efficient use of resources elsewhere. Evolution always involves trade-offs.
So, it was a very interesting article with exciting results and the media did not communicate it at all well because they couldn't get past the word bisexual. They didn't even check to see whether it was used to mean what they thought it meant. Again I am disappointed with the media's coverage of science.
Although I can't be sure, I think the churnalism all started with a story on Discovery News. The story does do a pretty good job of discussing the results of the study, but includes the sentence:
Male bottlenose dolphins also were found to engage in extensive bisexuality, combined with periods of exclusive homosexuality.This gets the situation completely wrong. An earlier paper2 showed that male dolphins in Shark Bay form alliances with one or two other males. These first order male alliances cooperate to aggressively herd females to gain mating access*. The first order alliances also prevent other males from mating with the females that they're herding.
The first order male alliances can also cooperate with other first order groupd to produce a second order alliance, which in turn may join to form a third order alliance. The higher level alliances cooperate to 'steal' females from smaller groups. But, lower level alliances can also recruit other first order groups to defend the female they are herding. If a female is successfully stolen, only one of the joined first level alliances will herd her afterwards.
The higher level alliances are much less stable than first order alliances. For example, groups A and B may cooperate to steal a female from group C one day, then groups B and C may cooperate to steal a female from group A on another day. However, some high level alliances may persist for well over a decade. The reasons for such fluid, reversible alliances at higher levels are unclear, but it may have to do with the context of the encounters.
Despite the findings being less 'juicy' than the popular science articles make it seem, the paper is really interesting. The purpose of the paper was to test hypotheses about the structure of dolphin societies. Evidence from previous studies, such as the one described above, is strongly suggestive that males form their strong alliances in otherwise open social networks. This type of social structure has never been previously documented in mammals.
In mammals, male alliances form to acquire or defend valuable resources from other males, such as territories or female groups. Although it did not seem like this was occurring in the Shark Bay dolphins, two hypotheses had not been properly tested. The 'community defence model', which argues that there are dominant alliances that range over the entire population, and the 'mating season defence model', which argues that males defend territories or females only during the breeding season.
The study found little evidence to support either of the two hypotheses. No male alliances ranged over the entire site as the 'community defence model' predicted. Alliances also overlapped during the mating season and females were not consistently associated with particular alliances as the 'mating season defence model' predicted. So, now we have strong evidence that dolphin societies are unlike any other mammal societies.
The authors of the study point out that a society organised in such a way would put unusual pressures on social cognition. Not only must they remember previous encounters with other dolphins, but they must cope with uncertainty that the relationships are still the same. Individuals that were friends in earlier encounters may have formed new alliances since you last interacted with them and become foes.
It's little wonder that social complexity correlates quite well with brain size, but it's hard to tell which came first. Larger brains may allow more complex social interactions to occur, but more complex social interactions may place selection pressure on large brain size. The authors argue that the complexity of dolphin society is probably a driver of brain size evolution in dolphins, and I tend to agree.
The authors also make the interesting argument that increased locomotion efficiency and low movements speeds have played a role in producing the social complexity and, therefore, brain size evolution. They argue that more efficient travel allows individuals to exploit larger areas and this would increase the chance that competitive encounters occur. A greater number of competitive encounters would, in turn, favour the formation of alliances with other individuals. I'm not so sure about this argument. Brains are energetically expensive to maintain, and probably favour more efficient use of resources elsewhere. Evolution always involves trade-offs.
So, it was a very interesting article with exciting results and the media did not communicate it at all well because they couldn't get past the word bisexual. They didn't even check to see whether it was used to mean what they thought it meant. Again I am disappointed with the media's coverage of science.
*It's been called 'gang rape' by some articles, but this is not at all what it actually is. Although, by human standards, it's definitely not romantic. The herding males threaten, bite, hit and crash into the females they are herding.
Further reading:
1 Randic, S., Connor, R. C., Sherwin, W. B., and Krutzen, M. (2012) A novel mammalian social structure in Indo-Pacific bottlenose dolphins (Tursiops sp.): complex male alliances in an open social network. Proceedings of the Royal Society B. doi: 10.1098/rspb.2012.0264
2 Connor, R. C., Smolker, R. A., and Richards, A. F. (1992) Two levels of alliance formation among male bottlenose dolphins (Tursiops sp.). Proceedings of the National Academy of Sciences 89: 987 - 990
Subscribe to:
Comments (Atom)